

Observer des régularités et définir des observables pertinentes dans l’étude des systèmes sociaux est une tâche ardue. Ce défi est l’objet des sciences humaines et sociales qui s’attèlent à en proposer des descriptions. Pour se faire, elles ont recours à diverses approches ; interprétatives, en se concentrant sur la manière dont les entités sociales (individu, groupe…) développent et interprètent des codes sociaux au travers des pratiques sociales (langage symbolique, rituel, etc. Et mécanistes, consistant à réduire les phénomènes sociaux à des catégories objectives, qui soient à la fois stables et générales.
Une telle diversité d’approches peut se comprendre par la variabilité inhérente aux systèmes sociaux ainsi que par la tension que cette variabilité peut générer avec une volonté scientifique d’identifier et de décrire des régularités. Cette tension émerge du fait que la singularité des objets tend à les rendre incomparables, voire incommensurables. Il est alors difficile de faire davantage que des études de cas et des reconstructions a posteriori. Par ailleurs, , nous pouvons avoir l’ambition de proposer des clés de compréhension qui permettent d’embrasser des cas en apparence singuliers sous des catégories plus générales, afin d’améliorer notre compréhension du social au-delà des cas passés et particuliers.
Cette tension s’illustre bien dans la cliodynamique, qui tente de caractériser de telles régularités en construisant des bases de données historiques et en s’appuyant sur de nouvelles méthodes comme la modélisation et l’analyse de données. Cela, afin de détecter sur le temps long et sur des échantillons très variés des régularités dans la manière dont les sociétés se transforment au cours du temps. Il est possible rapprocher cette nouvelle manière d’aborder les systèmes sociaux à des pratiques plus anciennes, cherchant à établir des critères de comparabilité généraux au sein des systèmes sociaux et de découvrir des mécanismes régissant leurs transformations. On peut alors situer cette approche dans une perspective dite « positiviste », c’est-à-dire dans la croyance qu’il existe des mécanismes universels, des «lois» du social au même titre que les lois de la physique et qu’elles n’ont simplement pas encore été découvertes. Cela aurait pour conséquence que les sciences sociales auraient vocation à tendre théoriquement et méthodologiquement vers les pratiques scientifiques de la physique.
Le programme positiviste semble cependant compromis par la nature même de l’objet d’étude des sciences sociales. Les objets sociaux, à l’instar des organismes biologiques, sont des entités ouvertes sur leur environnement dont le comportement n’est pas seulement le produit de causes pré-établies mais d’une interprétation émergeant de la friction entre organismes ayant une histoire et un environnement changeant.
Cette inter-action peut alors conduire à l’émergence de propriétés spécifiques, résultant de la résonance entre un état particulier de l’organisme, caractérisé par son organisation et son histoire, et celui de son environnement. De sorte que les deux apparaissent comme les deux faces d’une même pièce ; ils sont indissociables.
Dans cette relation d’interdépendance, ces propriétés spécifiques sont à l’origine de «nouveautés nouvelles», c’est-à-dire qui ne sont pas réductibles à une description a priori du système par des propriétés génériques (Montevil 2019). Pour reprendre l’analogie avec le langage développée dans un article précédent, l’interprétation n’est pas seulement le produit d’un « jeu de symbole » préexistant, mais de sa collision avec un contexte, une situation qui permet alors l’émergence de sens.
Cette situation crée une rupture radicale avec les méthodes de la physique, héritées de la dualité cartésienne sujet-objet, qui consistent à séparer objet et environnement, en négligeant certains phénomènes au profit d’autres. Ce geste fondateur qu’est l’acte de “négliger” se manifeste par la caractérisation d’un système isolé de son milieu par des abstractions, permettant d’établir des lois de conservation et la mise en équation de phénomènes ; en d’autre termes, l’établissement de sa description déterministe ou probabiliste.
En considérant les systèmes biologiques comme une interface entre les systèmes physiques et les systèmes sociaux, dans la mesure où l’on peut construire un environnement contrôlé pour des premiers tout en retrouvant les problématiques des seconds, on adopte ce que l’on nomme une approche « naturaliste ». On peut comprendre cette approche comme la supposition d’une continuité phénoménologique entre ces systèmes. Le naturalisme s’oppose ainsi aux conceptions prêtant des propriétés singulières aux systèmes sociaux comme le libre arbitre.
Dans cette perspective nous mettrons la focale sur la biologie en tant qu’interface épistémologique entre les sciences physiques et les sciences sociales. Nous verrons comment elle peut nous permettre de développer des outils généraux et pertinents pour traiter l’historicité (le rôle joué par l’histoire du système dans son évolution) et l’ouverture du système d’intérêt sur son environnement. Dans un premier temps, en comprenant le rôle du dispositif expérimental sur la variabilité des organismes. Puis, dans un second temps, la manière dont le découplage temporel entre des processus permet une forme de reconstruction « in vivo » du rôle joué par ces dispositifs expérimentaux. Enfin, nous développerons la perspective d’appliquer ces outils aux systèmes sociaux et les limites de cette extension.
Canaliser expérimentalement la variabilité
Dans une perspective naturaliste, la singularité des systèmes sociaux apparaît comme particulièrement saillante au regard des difficultés à les isoler de leur environnement et à construire des situations expérimentales contrôlées. A cet égard, l’étude des organismes biologiques semble plus accessible par ces méthodes expérimentales et semble donc être une interface pertinente pour étudier le rôle de l’historicité et de l’ouverture du système d’intérêt sur son environnement.
L’étude des organismes biologiques en laboratoire consiste en un ensemble de gestes et de dispositifs ayant pour objectif de les rendre comparable entre eux. Comme évoqué précédemment, les organismes biologiques sont tous le produit d’une histoire singulière (phylogénétique, généalogique et épigénétique pour ne citer qu’eux, et d’un environnement singulier. C’est ce que (Montevil et al 2016), nomme « principe de variation ».
Ainsi, les étudier nécessite tout d’abord de créer des catégories permettant de les comparer. C’est par exemple ce que nous faisons lorsque nous identifions des espèces. En adoptant une définition particulière du terme, nous pouvons dire qu’il s’agit de l’ensemble des organismes partageant la même histoire phylogénétique (évolutive).
Ces organismes deviennent alors interchangeables au sein de cette catégorie. L’intérêt porté à une espèce suppose donc que nous pouvons indistinctement étudier n’importe lequel de ses représentants. On peut de cette manière définir d’autres catégories à l’aide de la systématique comme le groupe, le clade ou des échantillons particuliers partageant des conditions communes.
La mise en laboratoire de ces organismes consiste donc à réduire leur variabilité inhérente afin d’établir des catégories, des symétries entre différents individus par des propriétés qu’ils partagent.
La notion de symétrie est ici importante et renvoie à une conception plus abstraite que celle de la symétrie géométrique, bien qu’elle soit dans les faits similaires. Par exemple, deux individus d’une même espèce sont symétriques (au yeux de la systématique, qui établit cette catégorie) au regard de leur histoire phylogénétique. En conséquence, on peut les interchanger dans la mesure où l’on étudie une caractéristique supposée dépendre principalement de leur phylogénie, c’est -à -dire de leur histoire évolutive comme on peut interchanger les deux figures géométriques sur leur axe de symétrie sans percevoir la différence.
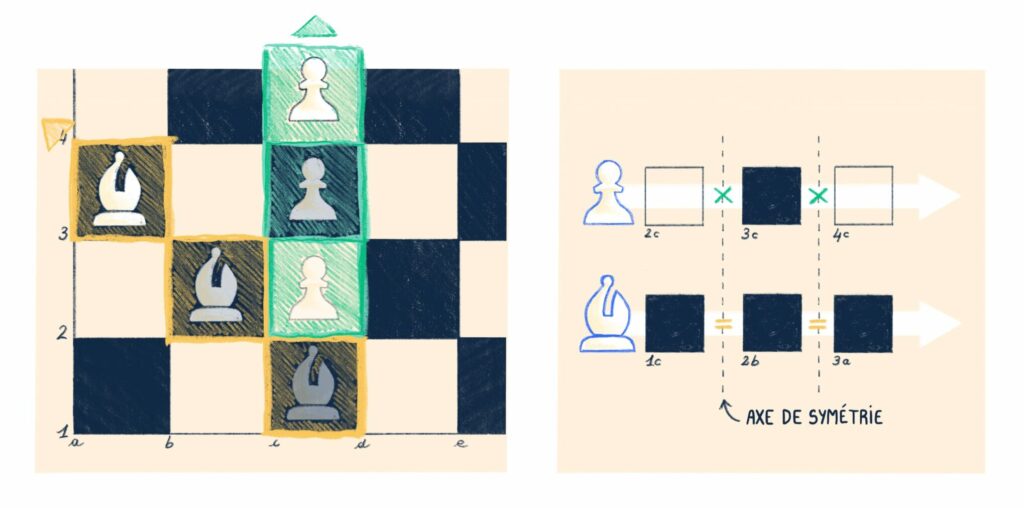
On peut étendre cette idée à afin de comprendre la catégorie d’élément chimique. Si l’on considère tous les atomes de l’univers comme des entités a priori unique produit par une histoire et des conditions singulières, nous pouvons comprendre le tableau des éléments de Mendeleiv comme une table de symétrie nous informant qu’au des processus chimiques, il est possible d’interchanger un certains nombre d’entre eux, ils forment des catégories ; ce sont les éléments. C’est-à-dire que l’on peut les permuter et sans modifier le cours d’un processus chimique. Tous les atomes d’hydrogènes partagent la propriété de contribuer identiquement aux réactions peu importe leur histoire. Ces catégories ne sont cependant définies qu’au regard des réactions chimiques, un romancier s’intéressant à compter l’épopée d’un atome d’hydrogène fera lui une différence entre eux, en fonction de leur histoire, ils ne seront donc plus permutable, la symétrie est rompu quand il s’agit de romancer (ou d’autres choses !
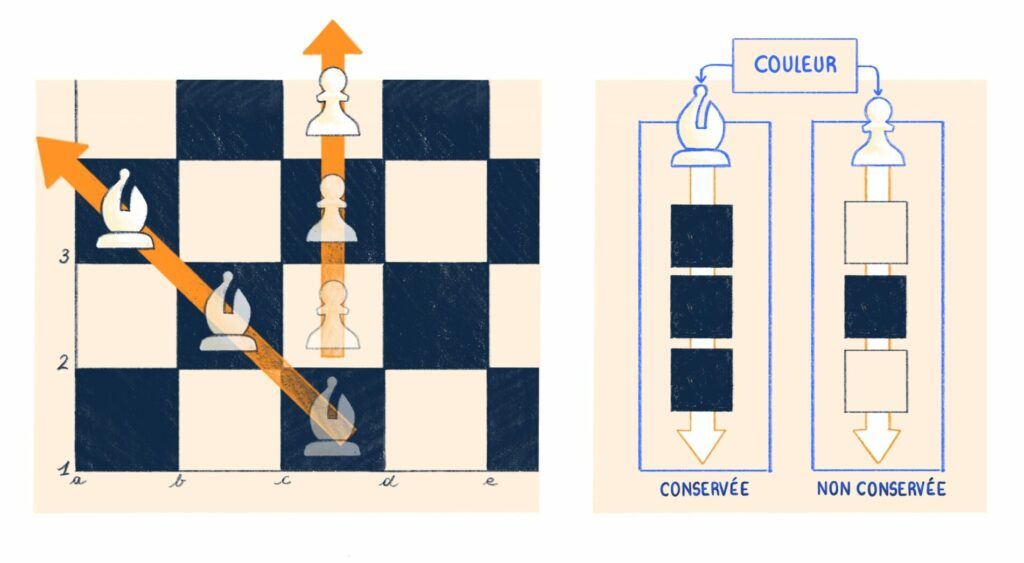
Revenons à la biologie. En poursuivant cette idée de symétrisation, on peut comprendre le laboratoire comme consistant à composer un ensemble de contraintes sur un échantillon biologique en vue d’étudier un phénomène précis. Ces contraintes peuvent permettre de contrôler la généalogie sur plusieurs générations, ou encore le développement des individus et l’évolution de leur environnement. Ce faisant, elles opèrent une symétrisation (on imposent des symétries) sur l’échantillon en induisant une histoire généalogique, épigénique ou un environnement commun.
La symétrisation induite par les dispositifs expérimentaux comme consiste à maintenir constants certains aspects de l’objet d’étude afin de réduire sa variabilité et ainsi pouvoir proposer des socles comparatifs. En somme, à définir des quantités ou des qualités conservées au cours du processus. On peut toutefois remarquer que cette constance n’est que relative, elle n’est assurée que sur l’intervalle de temps de l’expérience. C’est le fait que cette constance (ces symétries) joue un rôle dans le processus étudié qui lui permet d’agir comme une contrainte sur celui-ci.
Il apparaît cependant une limite à symétriser expérimentalement un échantillon Montevil (2019, 2). Plus on spécifie des conditions en créant des symétries, moins la portée des conclusions que l’on peut en tirer sera générale. Plus on augmente le nombre de conditions dans lesquelles la régularité étudiée peut être observée, moins il est vraisemblable de l’observer sans avoir à construire artificiellement ces conditions. Cette tendance ne fait toutefois pas office de lois, en témoignent les exemples suivants ; L’étude de cas, qui, tout en étant très peu spécifique (on prend un individu particulier sans imposer de symétrie) n’est pas plus générale. A contrario, l’étude de la fréquence cardiaque maximale (maximum basal rate) ne semble que peu dépendre de symétries à imposer, celle-ci dépendant principalement de l’histoire évolutive, ce qui constitue une condition que l’on retrouve sans contrôle expérimental.
Canaliser la variabilité in vivo
En comprenant les contraintes comme une affaire de rythme de changement relatif dans la production d’un processus qui en dépend, on peut élargir son application au-delà du laboratoire pour comprendre des processus « in vivo ». C’est ce qu’ont fait Maël Montevil et Matteo Mossio dans un article de 2015 intitulé « Biological organization as closure of constraints », dont le présent article s’est largement inspiré.
Dans cet article, les deux chercheurs proposent une description du phénomène d’auto-organisation en considérant les contraintes comme des causes efficientes. La différence entre causes matérielles (les flux de matières) et les causes efficientes (canalisations de ces flux) à été introduite par Aristote dans sa Physique, pour rendre compte de la différence entre la manière dont une chose est matériellement produite et les circonstances qui lui permettent de se produire. Lorsque je respire la cause matérielle est l’aspiration, mais la cause efficiente est la présence de poumons. Dans cet exemple, on a même une rétroaction entre processus et contrainte car ils sont entretenus grâce à l’oxygène respiré .
Ainsi une contrainte donnée est définie comme jouant un rôle actif dans la production d’un processus donné sans y contribuer par un transfert de matière. L’auto-organisation peut alors être comprise comme une rétro-action entre des causes matérielles et les conditions de leur réalisation : une rétro-action entre des contraintes et les processus qu’elles contraignent.
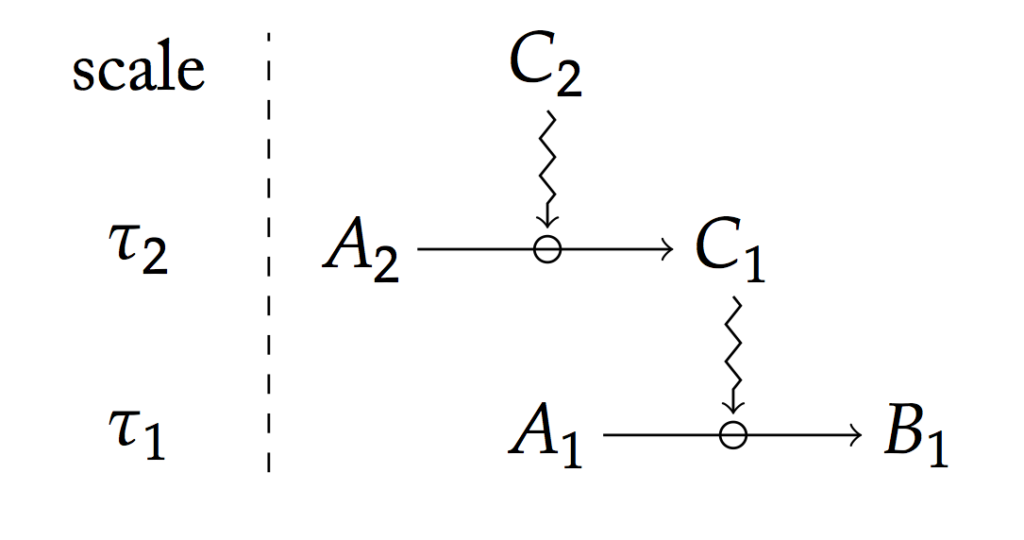
Cette distinction entre contraintes et processus n’est cependant pas de nature « ontologique » comme pourrait le laisser penser la distinction entre causes matérielles et causes efficientes. Une contrainte n’est définie que par rapport à un processus donné et dans la mesure où la contrainte qu’elle impose est induite par sa constance relative à l’échelle du processus. En d’autres termes, c’est parce que ce qui constitue matériellement la contrainte change plus lentement que le processus et que ce dernier en dépend pour sa réalisation, que cette dernière agit comme contrainte sur le processus.
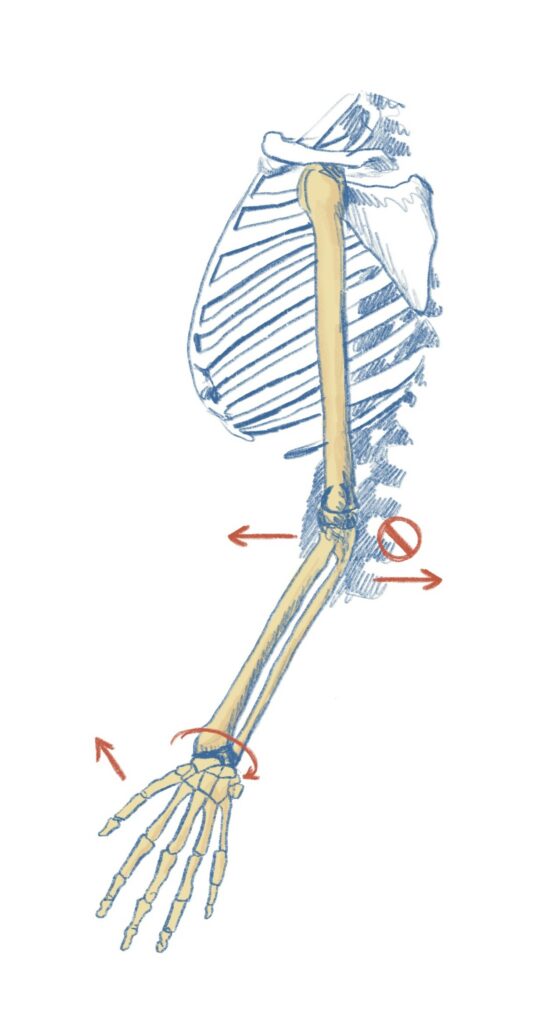
C’est parce que le développement et la variation matérielle des os varient plus lentement que le mouvement qu’ils permettent que ceux-ci agissent comme une contrainte sur le mouvement. On voit bien ainsi l’ambivalence du concept de contrainte ; elles permettent le processus tout autant qu’elle le contraigne et c’est d’ailleurs car elle le permet en réduisant la variabilité qu’elle le contraigne également.
Une des forces du concept de contrainte est aussi qu’il jouit d’une définition précise. Formellement, une contrainte est définie comme canalisant un processus par le fait de jouer un rôle dans sa réalisation tout en ayant un temps de variation plus important que celui-ci, c’est ce que représente le schéma ci-dessus. Il devient alors possible de composer les contraintes pour rendre compte de l’interaction entre différents processus ayant lieu dans un organisme (voir schémas ci-dessous).
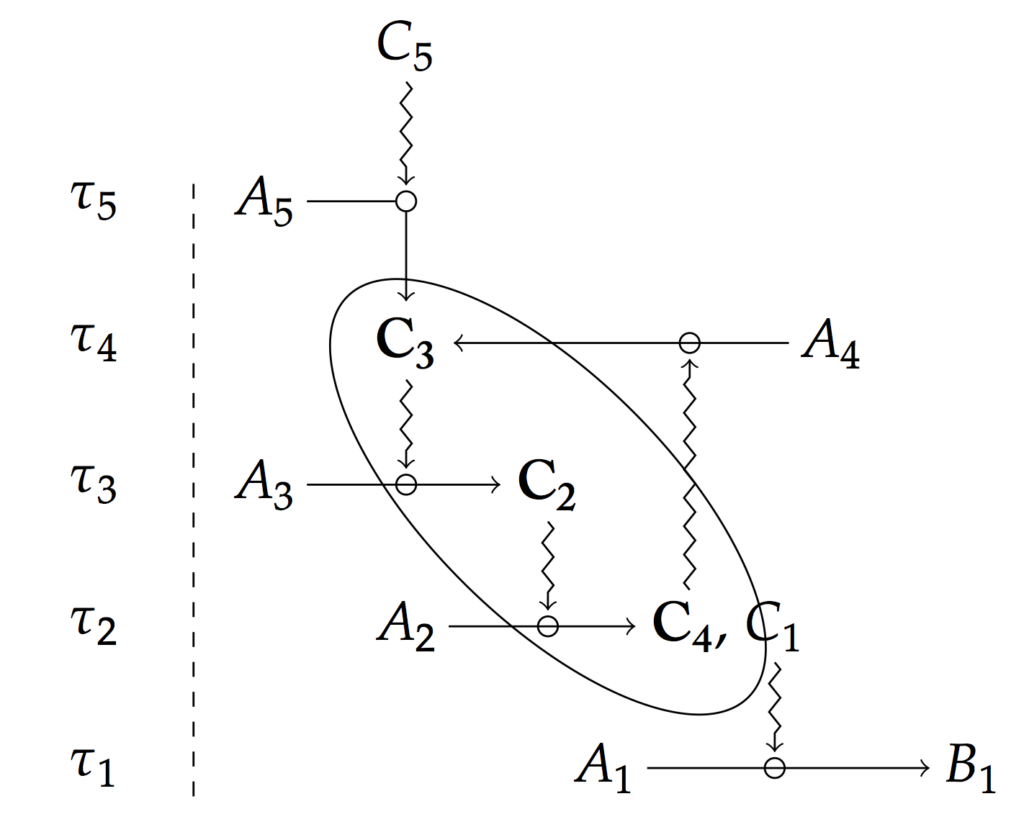
Le point clé de l’article n’est cependant pas tant de proposer une définition formelle du coept de contrainte que d’identifier les contraintes sujettes à une clotûre permettant de définir une fonctions biologiques, comme dans l’exemple précédent sur les poumons et la respiration.
Au delà de la biologie
Nous avons commencé cet article en mettant l’emphase sur les phénomènes d’historicité et d’ouverture comme source de variabilité communes aux systèmes biologiques et aux systèmes sociaux. Ce faisant, nous avons adopté une approche naturaliste supposant une continuité phénoménologique entre ces deux mondes et considéré les systèmes biologiques comme une interface pertinente pour l’étude de ces phénomènes.
Cela nous a permis de décrire la manière dont est gérée cette variabilité au sein des sciences biologiques au travers de pratiques expérimentales et des concepts généraux de symétrie et de contrainte. Ainsi, nous avons montré comment ces derniers pouvaient également être recrutés pour décrire in vivo, l’auto-organisation au sein des systèmes biologiques.
De part sa généralité et son usage courant, il peut être séduisant de recruter le concept de contrainte en dehors du cadre de la biologie dans la mesure où il semble reposer sur des considérations beaucoup plus générales comme les symétries et l’hétérogénéité des rythmes de changement. On peut par exemple y voir un usage en linguistique, avec la différence de rythme entre la langue et la parole, cette première pouvant faire penser à une contrainte de la seconde dans la mesure où elle la contraint autant qu’elle le permet, comme il a été abordé dans l’article « le social comme langage.
Cependant, il faut se garder d’étendre hâtivement les concepts de contrainte et de symétrie à l’ensemble des situations qui pourraient présenter des circonstances d’applications appropriées. Les contraintes sont actuellement définies dans un cadre théorique précis et propre à la biologie. L’étendre sans l’ancrer dans un autre cadre théorique spécifique pourrait aisément conduire à un mauvais usage du concept et à le vider de son contenu. Son emploi dans un cadre théorique précis est crucial car il permet de renvoyer à des situations empiriques précises, ce qui lui permet d’être sujet à des résultats négatifs et ainsi d’en juger la pertinence : c’est ce qui en constitue sa plus-value scientifique.
On peut toutefois noter la résonance du concept de contrainte dans la philosophie pragmatiste. En effet, une des caractéristique générale du concept tel que définie dans le cadre des systèmes biologiques est de reposer sur une hétéro-chronie des processus.
Or, la définition d’échelles de temps pertinentes dépend entièrement de l’objectif d’étude, si la relation entre contrainte et processus est préservée, l’identification des symétries et des contraintes pertinentes peut varier d’un objectif d’étude à un autre. Cela se comprend bien dans les différents dispositifs expérimentaux mis en place en vue de faire une observation spécifique en imposant un ensemble de symétrie spécifique.
Ainsi, la clotûre entre contrainte et processus, qui est pertinente à considérer, apparaît l’être au regard d’un horizon temporel donné. Si on élargissait le diagramme présenté plus haut, nous verrions d’autres clotûres apparaître, mais elles apparaîtraient au-delà de la fenêtre temporelle définie pour étudier un processus spécifique, observer une fonction spécifique. Cela illustre ainsi l’idée qu’il n’ y a pas d’observables absolus et a priori. Leur définition dépend d’un contexte théorique particulier, c’est-à-dire d’un usage spécifique traduit dans la situation expérimentale.
Nicolas Salerno, 2024
Remerciement à Emilie, Hélène, et Anton pour la relecture
Remerciement particulier à Hélène Bezin–Chaingy pour les illustrations
Références :
(Montevil 2019) : Maël Montévil. Possibility spaces and the notion of novelty: from music to biology, 2019.
(Montevil 2019b) : Maël Montévil. Measurement in biology is methodized by theory. 2019.
(Montevil & Mossio 2015) : Montévil, Maël, and Matteo Mossio. Biological Organisation as Closure of Constraints. 2015.
(Montevil & al 2016) : Montévil, Maël, Matteo Mossio, A. Pocheville, and G. Longo.Theoretical Principles for Biology: Variation. 016.